
Glycogen Supercompensation Enhances Athletic Performance
Michael C. Prevost
Aerospace Physiologist - Marine Aircraft Group 39, Camp Pendleton, CA.
Abstract
Glycogen supercompensation enhances athletic performance, Prevost, M.C., J. Perf. Enhan. 1999 1(1):1-10. The glycogen supercompensation effect (achieving supraphysiological glycogen levels due to carbohydrate depletion followed by loading) was first demonstrated in 1967. Since that time it has become a very popular pre-event performance enhancing strategy. Endurance athletes benefit from glycogen supercompensation because fatigue in events lasting longer than one hour is related primarily to glycogen depletion. Bodybuilders benefit from glycogen supercompensation because each gram of glycogen is stored with 3 grams of water; therefore a doubling in glycogen stores can increase the bodybuilder’s apparent muscle mass. Glycogen supercompensation is probably of little use to power athletes since fatigue in these events is not related to glycogen depletion and the weight gain may be a liability. Glycogen supercompensation occurs only when a low carbohydrate diet is combined with vigorous exercise followed by a high carbohydrate diet. Glycogen supercompensation occurs only in muscles that were trained and is maximal at a carbohydrate intake of approximately 25 grams per hour for average adults and possibly 40 grams or more per hour for bodybuilders.
Keywords: glycogen; carbohydrates; carbohydrate loading; glucose
Physiology Lesson
Carbohydrates are an important source of fuel for the body during physical activity and at rest. Because carbohydrates play such an important role in producing energy for the body, they are stored in the form of glycogen in both skeletal muscles and in the liver. Glycogen is essentially a long chain of glucose (sugar) molecules that are attached end to end with an occasional cross linkage. Figure 1 shows the basic structure of glycogen. Note that the glucose molecules are attached end to end. A glycogen molecule would consist of thousands of these linkages with occasional cross linkages as illustrated in Figure 2.
Figure 1. Glycogen Structure
Figure 2. Glycogen Structure With Cross Links

Glycogen makes up about 1-2% of muscle weight and 6-10% of liver weight. Although the liver has a higher concentration of glycogen than muscle there is more glycogen stored in muscle tissue because muscle tissue is more abundant than liver tissue. The average person would store about 400 grams of glycogen in their muscles and 100 grams in their liver. Since 1 gram of carbohydrate contains 4 Calories, the body stores approximately 2000 Calories in the form of muscle and liver glycogen.
The glycogen stored in muscle and liver comes from dietary carbohydrates if sufficient quantities of dietary carbohydrates are consumed. Once ingested, the carbohydrates that we eat are digested to simple sugars by pancreatic and salivary amylase. Sugars other than glucose are largely converted to glucose in the small intestine. Some sugars like fructose (the primary sugar in fruits) when consumed in significant amounts (the amount varies but for fructose it is typically 50g or more per meal) may enter the bloodstream in their native form and must be converted to glucose by the liver. Once the sugars reach the bloodstream they are driven into the liver and muscle cells by the action of the hormone insulin. Insulin also activates the enzyme glycogen synthase, which synthesizes glycogen by adding one glucose at a time to the glycogen chain. When muscle and liver glycogen levels are replenished, the excess glucose is burned preferentially to fat. In other words, consuming excess carbohydrates (more than is needed to replenish glycogen stores) shifts the body from burning primarily fats at rest to consuming primarily carbohydrates at rest. This process is also activated by insulin. If caloric intake is in excess of caloric need, the excess carbohydrates will be stored as fat.
The body cannot use glycogen as a source of energy in its storage form. In order to derive energy from glycogen the body must liberate individual glucose molecules to use for energy production. This process is accomplished by activating the enzyme glycogen phosphorylase. Glycogen phosphorylase removes glucose units, one at a time, from the glycogen chain. Glycogen phosphorylase is activated automatically when muscles begin to contract. When muscles contract, the calcium concentration inside of the muscle cell increases. Calcium converts the inactive form of glycogen phosphorylase to a more active form of glycogen phosphorylase. This provides glucose to fuel the muscular activity.
If the exercise continues for any length of time the body begins to produce hormones that also activate glycogen phosphorylase. Some of the hormones are glucagon, epinephrine and norepinephrine. These hormones activate both muscle and liver glycogen phosphorylase. Although the liver is metabolically active, most of the glucose that is produced is released into the bloodstream. The glucose that is released by skeletal muscles is trapped inside of the muscle cell and cannot contribute to blood sugar. The liver release of glucose from glycogen helps to maintain blood sugar levels during exercise. Some of the blood sugar is used as a source of fuel by the nervous system and some is taken up by skeletal muscle as an alternate source of glucose to fuel muscle contraction.
Review of the Literature
Muscle glycogen may be the most important energy substrate during exercise. At the beginning of all types of exercise and for the entire duration of high intensity exercise, muscle glycogen serves as the primary metabolic energy substrate (1Powers and Howley, 1990). Because muscle glycogen concentration influences endurance performance (2 Conlee, 1987) and may also affect maximum power output (3 Heigenhauser, Sutton and Jones, 1983), manipulating glycogen stores is a potentially important consideration for a wide variety of athletes.
In 1967 Ahlborg and colleagues (4) began to demonstrate the relationship between diet and muscle glycogen concentrations. Figure 3 below demonstrates the results of the study. When subjects consumed a low carbohydrate diet glycogen concentrations decreased then rebounded to double baseline concentration on a high carbohydrate diet. This effect (increased glycogen storage ability following glycogen depletion when consuming a high carbohydrate diet) is referred to as glycogen supercompensation. Glycogen supercompensation is different than glycogen compensation. Glycogen compensation is a normal response to exercise and refers to the process of replacing muscle glycogen to normal levels following exercise. Supercompensation occurs when glycogen concentrations are replaced to supra-physiological levels (much greater than normal). A more recent study (5 Goforth, Arnall, Bennett and Law, 1997) demonstrated that glycogen concentrations remain supercompensated (elevated over baseline) for at least 3 days following a carbohydrate depletion/loading cycle. Since the glucose that is produced from muscle glycogen does not leave the muscle cell, glycogen is only depleted in muscles that are exercised (6 Hultman, 1971). The amount and type of carbohydrate ingestion that will maximize glycogen resynthesis has been the subject of numerous research studies. Costil et al. (7) demonstrated that the rate of glycogen resynthesis was greatest when carbohydrate intake was 525 grams per day or more. Blom et al (8) showed that glycogen resynthesis was maximal when subjects consumed 25 grams of glucose per hour. Keizer et al. (9) showed no further increase in the rate of glycogen synthesis measured by Blom at an intake of 70 grams of carbohydrates per hour. Based on these studies it appears that 25 grams of carbohydrates per hour (600 grams per day) is sufficient for a maximal rate of glycogen resynthesis. A study by Roberts et al. (10) demonstrated that glycogen resynthesis was greatest in subjects who consumed simple sugars rather than complex carbohydrates. The simple sugar fructose (the primary sugar found in fruits) is effective at replenishing liver but not muscle glycogen (11). This is because muscle tissue lacks the enzyme necessary to convert fructose to glucose. Therefore fruit is a bad carbohydrate choice for carbohydrate loading or supercompensation.
Endurance Athletes
One of the earliest studies on the effects of muscle glycogen on endurance was conducted in 1967 by Ahlborg and colleagues(4). In this now classic study, he demonstrated a correlation between initial muscle glycogen concentration in the vastus lateralis muscle of the quadriceps and exercise endurance using a continuous bicycle ergometer protocol. Since this study there have been numerous studies validating this effect (see Conlee, 1987 for a review). In another classic study Bergstrom and associates (12) studied the effects of altering carbohydrate consumption for 3 days on exercise to exhaustion. The researchers had the same subjects consume a mixed diet, a high low carbohydrate diet, and a high carbohydrate diet for three days. After each three-day period glycogen concentrations were measured and the subjects exercised to exhaustion on a bicycle ergometer. The results are summarized in Figure 3.
Figure 3. Effect of Diet on Muscle Glycogen and Performance
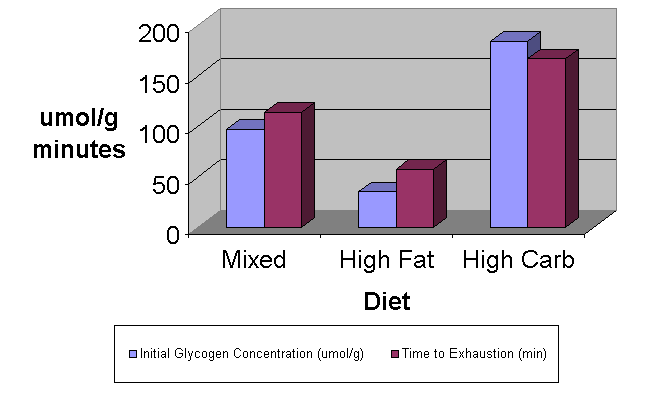
Note the dramatic differences in glycogen concentrations (Glycogen concentration is plotted on the y axis in umol/g.) and performance (Performance is also plotted on the y axis in minutes.) from only 3 days of dietary manipulations. The order of the treatments was mixed diet, followed by high fat – high protein and finally high carbohydrate. Not only did the high carbohydrate diet replace the carbohydrate stores that were depleted by the high fat – high protein diet, but it actually increased glycogen concentrations over baseline levels. Bergstrom and colleagues concluded that the ability to sustain prolonged exercise depends on muscle glycogen concentration. The myriad of studies that followed firmly established the theory that sustaining performance in endurance events lasting longer than one hour is strongly dependent upon maintaining glycogen concentrations and that fatigue during these events is probably due to glycogen depletion (2).
Power Athletes
Although glycogen depletion does not cause fatigue during high power events (13), glycogen depletion has been shown to reduce the ability to produce a high power output. Heigenhauser and associates (3) demonstrated that maximal power output was reduced by 14% on a standard power test on the bicycle ergometer following glycogen depletion. A standard power test involves pedaling as fast as possible against a fixed resistance for 30 seconds. Conlee (4) speculates that this reduction in power output occurs because some fibers are no longer capable of contributing because they are almost completely devoid of glycogen. Since there are fewer fibers available to contribute, power output is reduced. In support of this theory, Jacobs (14) found that subjects could perform 10-20% more work during 50 maximal muscle contractions with carbohydrate loading.
Bodybuilders
Since every gram of glycogen is stored with approximately 3 grams of water (13) a doubling of glycogen stores due to glycogen supercompensation is likely to increase the apparent size of muscles. Since exercise upregulates the body’s ability to store glycogen and bodybuilders have more muscle mass than the average person, we might expect that a bodybuilder stores considerably more than the 500 grams of glycogen mentioned earlier as an average value for normal adults. For the sake of argument let’s assume that a bodybuilder is storing 800 grams (not an unreasonable amount) of muscle glycogen. By carbohydrate depletion and supercompensation to twice that level (again, not unreasonable) it would be possible to add 800 grams of glycogen plus 2400 grams of water to the bodybuilder’s muscle tissue. This amounts to a 7.2 pound weight gain from a single carbohydrate depletion/carbohydrate loading cycle. Therefore a bodybuilder can potentially gain a significant amount of apparent mass with successful glycogen supercompensation.
Recommendations and Conclusions
Endurance Athlete
Beginning a typical 3-day depletion, 3 day loading supercompensation cycle just prior to a competition may not be the best strategy for an endurance athlete. This is because glycogen depletion requires vigorous exercise and most endurance athletes refrain from vigorous exercise during the final week prior to a competition to ensure adequate recovery. Fortunately glycogen levels stay elevated for at least 3 days following a glycogen supercompensation cycle (5). This allows the athlete to start the cycle 9 days prior to competition and still allow 6 days of recovery before the event. A typical glycogen supercompensation cycle would look something like this:
Table 1. Recommendations for Endurance Athletes
| Day | Diet | Exercise |
| 1 | High protein/low CHO | Vigorous |
| 2 | High protein/low CHO | Vigorous |
| 3 | High protein/low CHO | Vigorous |
| 4 | High CHO (600 g) | None |
| 5 | High CHO (600 g) | None |
| 6 | High CHO (600 g) | None |
| 7 | Normal | None |
| 8 | Normal | None |
| 9 | Normal | None |
*CHO = carbohydrate
The vigorous exercise should use the same muscles that are going to be used during the competition since it is these muscles that will be depleted and supercompensated. In other words, if you are a runner you carbohydrate deplete by running. If you are a cyclist you carbohydrate deplete by cycling. Most of the carbohydrate consumption on day 1 of the high carbohydrate phase should be simple sugars and intake should not exceed 25 grams per hour or 75 grams every three hours. Carbohydrates should be consumed at least every three hours so that continual glycogen synthesis is occurring.
Power Athlete
If, as Conlee speculated (4), some muscle fibers are completely glycogen depleted by high power performances and subsequently are incapable of contributing, one might speculate that power athletes could benefit by glycogen supercompensation. For many athletes, however, actual performance during competition would not be enhanced by supraphysiological levels of glycogen. For weightlifters, for example, performance is related to the ability to produce force and not the ability to maintain force output over time. Although glycogen loading can delay the reduction in force output during repeated maximal contractions (14), no study to date has shown that maximal force production can be enhanced by supraphysiological concentrations of glycogen. The same logic applies to jumpers and throwers. For high power events lasting less than 10 seconds (100m sprint) the majority of the energy comes from stored Adenosine Triphosphate and Creatine Phosphate with little contribution from carbohydrates (Brooks and Fahey 1987). For high power events lasting longer than 2 minutes performance is limited by the cardiovascular system (13). Based on these facts and the Heighenhauser study mentioned earlier (30 seconds of maximal pedaling), one might speculate that glycogen supercompensation might be useful for high power events lasting between 10 seconds and two minutes. However, there is an important distinction between power tests and other 30 second events like a 300m dash. In a power test power peaks early because subjects are pedaling maximally from the start. During all but the shortest sprinting events there is some degree of pacing. It is not known if pacing would affect the relationship of glycogen to fatigue during these events. In addition, no study to date has shown an actual increase in performance in sprinting events (either bike, run or swim sprints) due to glycogen supercompensation. Also, in some power events, like weightlifting and sprinting, extra bodyweight can be a liability. Although they should maintain an adequate carbohydrate intake to prevent a decrement in performance, there is no strong evidence to suggest that power athletes would benefit from glycogen supercompensation prior to competition.
Since training can involve repeated high power performances (repeated sprints, or sets) one might speculate that glycogen supercompensation might be an effective training aid. While training performance might benefit from high concentrations of muscle glycogen, athletes cannot glycogen deplete and supercompensate prior to every training session. Most athletes would be averse to performing a glycogen depletion/supercompensation cycle frequently because it is so unpleasant.
Bodybuilders
An apparent increase in muscle mass is certainly a bonus for bodybuilders. Therefore, successfully glycogen supercompensating can certainly be a worthwhile process for these athletes. Since bodybuilders have much more muscle mass than the average person, larger carbohydrate intakes are likely to be required to maximize glycogen synthesis. Therefore, instead of the 25 grams per hour mentioned earlier, a large bodybuilder is likely to need closer to 40 grams (This is assuming that the bodybuilder has 40% more muscle than the average person). Since we are trying to maximize glycogen supercompensation in all muscles, we must glycogen deplete all muscles. This is accomplished by performing high repetition, high volume workouts for all body parts while on a low carbohydrate diet prior to glycogen loading. A typical regimen might look like this:
Table 2. Recommendations for Bodybuilders
| Day | Diet | Exercise |
| 1 | High protein/low CHO | Vigorous |
| 2 | High protein/low CHO | Vigorous |
| 3 | High protein/low carb. | Vigorous |
| 4 | High CHO (.43g/kg/hr) | None |
| 5 | High CHO (.43g/kg/hr) | None |
| 6 | High CHO (.43g/kg/hr) | None |
The bodybuilder should be training the entire body over the three-day period with a large volume of high repetition exercises to enhance glycogen depletion. It is the total volume of work that will determine the degree of glycogen depletion so rest between sets should be adequate to allow a large volume of work to be performed. Bodybuilders should avoid lifting very heavy as high force eccentric contractions have been shown to interfere with glycogen synthesis (15) probably due to muscle microdamage. Additionally Doyle et al. (16) found that glycogen synthesis was 25% lower in muscle that had performed eccentric contractions 48 hours earlier. Although the bodybuilder might not normally train three days in a row, it is recommended in this case. This prevents the bodybuilder from having to remain on a low carbohydrate diet for more than three days. Determining the amount of carbohydrates that should be consumed will require some trial and error but the research literature might provide some clues. A study by Pascoe et al. (17) found that the glycogen synthesis rate following resistance exercise was 12.9 mmol/kg/hr. If you know the molecular weight of glucose and can convert mmol to grams and if we assume that each gram of glycogen is stored with 3 grams of water this would give us a value of approximately .43g/kg/hr (explaining the calculations is beyond the scope of this review). If we match carbohydrate intake to the glycogen synthesis rate this would equal 43 grams per hour for a 220 pound bodybuilder (100kg) and a total of approximately 1000g (4000 Calories) from carbohydrates in a 24 hour period. Ivy (11) recommends consuming a 1g/kg bolus (single feeding) immediately after exercise and adding protein to enhance the insulin response. Glycogen replenishment is very rapid for six hours after high intensity exercise (11) and glycogen concentrations can return to baseline levels within this six hour period if adequate carbohydrates are consumed (supercompensation occurs in the days that follow). Therefore providing a bolus as Ivy suggested might speed up the process relative to consuming a predetermined number of grams every 3 hours. On day 1 most of the carbohydrates should be in the form of simple sugars to enhance glycogen uptake. The degree of glycogen supercompensation can be estimated by the amount of weight gain. Recall that each gram of glycogen is stored with 3 grams of water. If a bodybuilder gained 1600 grams (3.5 pounds) then an extra 400 grams of glycogen was stored.
In summary, glycogen supercompensation can be a valuable performance-enhancing tool for bodybuilders and endurance athletes. However, there is no convincing evidence to recommend its use to power athletes.
References
1. Powers, S, Howley, E, Exercise physiology: Theory and application for fitness and performance, Dubuque, Iowa: Wm. C. Brown Inc., 1990.
2. Conlee S, Muscle glycogen and exercise endurance - a twenty year perspective. Exercise and sports science reviews 1987:15: 1-28.
3. Heigenhauser G, Sutton J, Jones N. Effect of glycogen depletion on the ventilatory response to exercise. J. Appl. Physiol. 1983 54:470-474.
4. Ahlborg G, Bergstrom J, Edelund G, Hultman E. Muscle glycogen and muscle electrolytes during prolonged physical exercise. Acta Physiol. Scand. 1967 70:129-142.
5. Goforth, Arnall D, Bennett B, Law P. Persistence of supercompensated muscle glycogen in trained subjects after carbohydrate loading. J Appl Physiol 1997 82(1):324-347.
6. Hultman E Nilsson H. Liver glycogen in man. Effect of different diets and muscular exercise. In: Muscle Metabolism during Exercise. New York: Plenum, 1971.
7. Costill D, Sherman W, Gind C, Maresh C, Witten M, Miller J. The role of dietary carbohydrate in muscle glycogen resynthesis after strenuous exercise. American Journal of Clinical Nutrition 1981 34:1831-1836.
8. Blom P, Hostmark A, Baage O, Kardel K, Machlum S. Effect of different post-exercise sugar diets on the rate of muscle glycogen synthesis. Medicine and Science in Sports and Exercise 1987 19:491-496.
9. Keizer H, Kuipers H, Van Kranenburg G, Geurten P. Influence of liquid and solid meals on muscle glycogen resynthesis, plasma fuel hormone response and maximal physical working capacity. International Journal of Sports Medicine 1987 8: 99-104.
10. Roberts A, Noble D, Hayden D, Talyor A. Simple and complex carbohydrate-rich diets and muscle glycogen content of marathon runners. European Journal of Applied Physiology 1988 57:70-74.
11. Ivy J. Glycogen resynthesis after exercise: effect of carbohydrate intake. Int. J. Sports. Med. 1998 19 (2): S142-5
12. Bergstrom J, Hermansen E, Hultman E, Saltin B. Diet, muscle glycogen and physical performance. Acta Physiol. Scand. 1967 71:140-150.
13. Brooks G, Fahey T. Exercise Physiology. New York: Macmillan Publishing Co. 1984.
14. Jacobs I. Influence of carbohydrate stores on maximal human power output. In: Exercise Benefits, Limits and Adaptations. London: E & F N Spon Limited, 1987.
15. Widrick J, Costill D, McConell G, Anderson D, Pearson D, Zachwieja J. Time course of glycogen accumulation after eccentric exercise. J Appl Physiol 1992 72(5):1999-2004
16. Doyle J, Sherman W, Strauss R Effects of eccentric and concentric exercise on muscle glycogen replenishment. J. Appl. Physiol. 1993 74(4):1848-55.
17. Pascoe D, Costill D, Fink W, Robergs R, and Zachaweija J. Glycogen resynthesis in skeletal muscle following resistive exercise. Med. Sci, Sports Exerc. 1993 25(3): 349-354.
Questions
1. Each gram of glycogen is stored with ____ grams of water.
2. Glycogen is essentially a long chain of _______ molecules.
3. Glycogen resynthesis averages _______ grams per hour for average adults.
4. The enzyme that breaks down glycogen is called ________.
5. T F Research indicates that by increasing carbohydrate intake from 25 to 70 grams per hour the rate of glycogen resynthesis can be increased.
6. T F During exercise, glycogen depletion only occurs in muscles that are exercised.
7. T F Glycogen concentrations return to normal 2 days following glycogen supercompensation.