EXTENDED ABSTRACT
It is proposed that the deciduous climax forests of North America are primarily a function of winter glaze ice, with shallow roots in swamp soil an auxiliary factor south of the glaze ice zone because of winter uprooting. The deciduous understory is proposed as resulting from inability to photosynthesize faster than herbivorous destruction while soils are frozen. The trees and their saplings are devoid of leaves, so that any few remaining under story evergreen plants would have to withstand severe grazing with little concomitant advantage from photosynthesis. Because of deciduous habit's ubiquity in angiosperms, it is likely that deciduousness arose well before the Cretaceous when they were first seen on the fossil record. Some features may have arisen in the carboniferous, possibly culminating in an unknown tropical continent ( the Ontong-Java Plateau?) in the vicinity of the western Pacific, probably in Permian time. Angiosperms were thus pre adapted to colonize northern Hemisphere glaze ice areas in the Tertiary, The dominant trees evolved from subtropical genera.
INTRODUCTION
Glaze ice is caused by warm moist air riding up over an air mass which is below freezing. As rain falls through the cold air it becomes super cooled and turns to ice a upon striking an object which is itself below freezing.
Evergreen trees, usually represented by the conifers, are the dominant upper story vegetation in the temperate, north temperate, and boreal regions, both in size of upland plants and area covered. In North America, they cover 3/5 of the forested areas and make up to 99.9% of the biomass in some areas of cool upland [Waring & Franklin]. As nearly as can be told, they have held this dominance far back in time (Douglas).
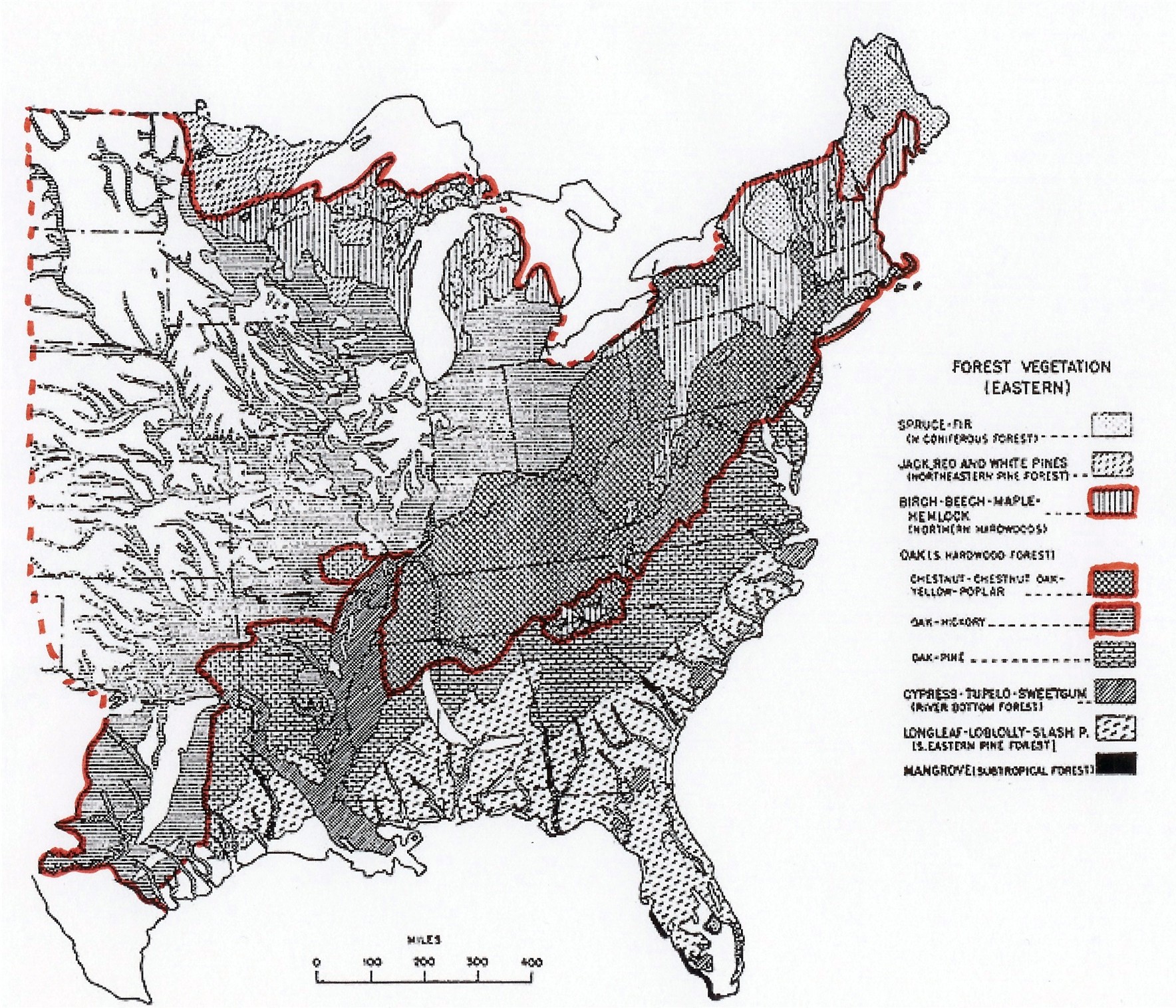
The boundaries between evergreen and the deciduous areas is approximately coexistent with the boundaries of the glaze ice areas. The maps show the close proximity of the boundaries. The forest map below, is from the U.S. Forest Service. The red line outlines the extent of areas that, except for a few patches on mountain tops, etc., within which the trees are almost pure deciduous stands.
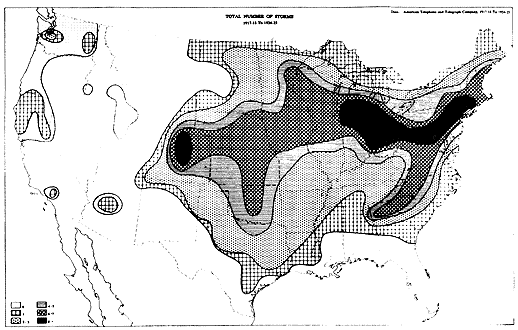
The glaze ice map below is from the A&T company. It is possible that if we had records for the last several hundred years the correlation would be even closer;
The western coniferous forests are not shown because, while there is somewhat of an association of deciduous vegetation with the small glaze ice areas and the riparian areas (author's observation), they don't show up on Shantz and Zon's adapted map.
According to Lemon, an ice storm will hasten the succession of the dominant trees [Lemon]. The loss of woody material from a major ice storm is close to the loss from the worst hurricanes. When attempting to explain structural variations in vegetation it is logical to search for mechanical causes. Of course non mechanical parameters such as nutrients, fire, rainfall, light, and temperature can have indirect and peripheral influences, but primary causes are likely to be mechanical. Deciduousness itself tends to be in an intermediate position, and as shall be developed, appears to be strongly influenced by grazing and soil water as well as glaze ice.
DISCUSSION
Since conifers furnish such effective competition to angiosperms outside the glaze ice zone, it is desirable to explain why any angiosperms at all can compete, especially those which lose leaves every year, for evergreens keep their leaves from 7 months to 4 years in the northern Amazonian rain forest [Sugden]. On the northern glaze ice border near Michigan, and to the north, the additional reinforcement provided by wet snow loads to glaze ice pressure may be an important reason why deciduous trees are still represented north of the glaze ice, although an increasingly smaller fraction of the trees [Braun p339]. I have noted myself that evergreen angiosperm vines are virtually nonexistent.
In the south, the deciduous canopy extends outside the glaze ice area, but only to any extent in the swamps and on the river bottoms. One can easily see the rather sharp demarcation in winter of standing water and its close association with deciduous defoliation if viewed from the air flying through Georgia and northern Florida [Monk, first Monk reference]. This last deciduous distribution may be due to shallow roots in soft, unfrozen soil coupled with winter winds, some of which can reach hurricane force. Reinforcing the concept that shallow roots may be determining is the fact that inside the glaze ice zone {in New Jersey}, I take note that swamp trees lose their leaves sooner than adjacent upland trees except for the shrubs and some shrub sized monocotyledons.
In the Northwest, it is possible that protection from the freezing of shallow mountain soils by early snow cover under the coniferous larches (larix) [Fosberg] is the reason why the pioneer larches are deciduous and thus avoid being blown over [Touzet]. The branches may be weak but the roots are tough and as deep as possible and thus assist in preventing blow downs. Further assisting them is the ability to straighten after being bent by snow loads.
The only deciduous angiosperm trees in this region, Montana, grow in swamps [Fosberg] [Nimlos] and river bottoms. The reason given for larches may be the main reason why deciduous trees grow up to the tree line only where there is winter snow in Norway. The Europeans, however, believe it is because the snow keeps the soil frozen in early spring. An additional indication that swamps place pressure on plants to become deciduous in temperate climates is that other orders have become deciduous in the past, such as Lepidodendron species in lower Pennsylvanian swamps [Michelle, et al].
The presence of patches of willow, alder, and aspen in the Arctic has been attributed to breakage of conifer branches by snow loads [Pruitt]. However I suspect that the reason the gap is not filled by conifers at first is winter grazing of seedlings by under snow herbivores. Herbivorous pressure must be a factor in the shrub layer in the heavy glaze ice zone [Fretwell] where all the shrubs are deciduous, even though they are not broken or uprooted much by ice [Downs]. Even the shrubs under the pine forests of northern Minnesota are hardwood [Braun, p387]. Shrubs under stands of Hemlocks were extensively Yews [Taxus canadiensis] but in recent years deer have destroyed them [Braun pp 362, 365] thus reinforcing this concept. If the trees and their saplings, which usually make up the bulk of the vegetation, are determined by the glaze ice, the small amount of shrub vegetation which would be left would be subject to intense pressure from grazing animals if they were evergreen. Middle aged trees are most damaged by ice [Illick], but very young trees which can be reached by herbivores require parents which are reasonably nearby. Apparently dominant trees have had no advantage evolving forms which are evergreen only in their youth. There are none, other than the larch, the leaves on the seedlings and saplings of which are protected from winter dehydration by snow cover [Richards] and possibly also from herbivorous pressure. However the larch does not grow inside the heavy glaze ice zone, which zone does not have constant snow cover.
Giving support to the concept that grazing pressure is an important determinant of understory deciduous habit is the circumstance that in savanna angiosperm forests in the vicinity of Sonora, west Texas, I take note that the large trees are almost all evergreen, while the small trees are predominantly deciduous. Sonora is outside the glaze ice zone. The growth is sufficiently open that the small trees are not understory.
On the southern swamp soils of southern Georgia, on the other hand, where the trees are predominantly deciduous conifers and deciduous angiosperms, I take note that the shrubs are primarily evergreen. The mild winters and adequate moisture not compromised by frost penetration of the soil are probably the reason why low plants gain an advantage from winter photosynthesis while enduring winter herbivores and are successful. Winter may be the only time when these shrubs receive adequate light. Evergreen angiosperm trees are somewhat more successful on southern upland soils than in the west, such as the live oaks {Quercus virginiana}, but conifers predominate except near the Gulf Coast [Braun p303]. The live oaks are really deciduous, but they suddenly lose all their leaves in the spring so the effect is evergreen. Deciduous trees are only a small part of upland trees south of glaze ice. An exception to conifers is the increasing dominance of evergreen magnolia {Magnolia grandiflora} southward on the fertile calcareous soils supporting the Beech-Magnolia climax [Braun p307]. In the vicinity of Chattanooga, Tennessee there is a sharp demarcation between deciduous on northwest slopes and evergreen on southeast slopes. I do not know if there is a glaze ice reason for such a sharp line such as warmer branches on southeast slopes, but it would seem to invalidate the fire climax hypothesis since any fire would surely not stop suddenly at the ridge crest.
It is also in order to consider the converse of the above; why evergreen conifers, which are so susceptible to glaze ice damage and therefore must make such disadvantageous compromises in competing for light, survive inside the glaze ice zone. In southern New Jersey, Maryland and Virginia mixed stands of pine and oak exist just inside the glaze ice zone. However, they grow on sandy soils with poor moisture retention and fertility. As a result, the vegetation is low and sparse. Compromises for light gathering to be discussed later are not important and losing nutrients in falling leaves is important [Aerts] [Monk, second reference]. Where tall luxuriant stands exist, the deciduous trees predominate. Stands of conifers grow on Appalachian peaks inside the glaze ice zone. Two air masses can not exist over these peaks at once so the glaze storms can not occur. As a result the worse storms occur below 1200 meters [Abell]. Below those altitudes, hemlock {Tsuga canadiensis} is a part of the moderate ice zone from Michigan through Pennsylvania to Massachusetts. Hemlock are said to be characteristically trees of ravines in the south [Braun p68], rocky talus slopes [Braun p102], or associated with dry soils [Braun p65]. Eastern red cedar {Juniperous virginiana) is common in New Jersey, but only on soils which have been so badly eroded by past farming that few other trees grow and therefore light is not limiting. Thus their compact growth is not a disadvantage. Sandy acid Silurian derived soils may account for the few pitch pines which grow on ridges in northwestern New Jersey [Niering]. Sandy soils may also account for the patches of white pine which grow in the Great Lakes area [Maycock] near the edge of the glaze ice zone. In Colorado and Wyoming stands of ponderosa pine {Pinus ponderosa} and blue spruce {Pica pungens} exist inside the glaze ice zone. These are probably a function of a short growing season combined with dry weather and light icing. If summers are dry, winter photosynthesis becomes more important and conifers are able to synthesize even below freezing [Waring and Franklin]. In the Columbia River gorge there is a zone of icing which strips the branches off the conifers on one side only [Lawrence]. Perhaps the fact that part of the tree remains relatively immune is part of the reason why so many conifers are able to grow there.
Some species of oak trees retain their dead leaves through the winter. Such trees are fairly common in Minnesota. However they do so primarily on the lower branches of the south side which are kept short, and have a further advantage of allowing the dead leaves to droop down as a result of which the leaves shed much of the water before freezing. I suspect that this gives them the advantage of protecting the bark of the main trunk somewhat from the stress of solar insolation cracking on very cold days without endangering the main limbs from ice.
It is not possible to invoke other meteorological parameters to explain temperate deciduous distribution. Winter isotherms run almost perfectly parallel to latitude in North America, and the few deviations from this show no relation at all to the deciduous zone. Rainfall isopleths {lines of equal depth} tend to parallel longitude, and in any case there are extensive evergreen stands in all possible rainfall districts elsewhere. Wind has already been suggested as playing a role on swamp soils, but on most upland soils winter wind is not strong enough to cause extensive uprooting in the absence of ice. In the section where glaze ice is heaviest, I take note that oaks, beeches, hickories and maples which have asymmetrical branches growing out at large upward angles to the trunk {often more than 60 degrees}, no leaves in winter to catch freezing rain, and extremely strong dense wood which permits long flexible branches are the dominant upper story trees and make up well over half the trees. So characteristic is strong dense wood of this region that they are often referred to as "hardwood forests". The only evergreens of consequence are the occasional stands of hemlocks and white pines mentioned previously. If one removes unusually sandy, acid, shallow, or infertile soil from the delineation, the displacement from heavy ice areas of evergreen trees is excellent, almost complete.
Glaze ice favors trees which have winter leaf fall (deciduous), short branches, flexible wood, tough hard wood, down sloping branches, and branches which do not make a small upward angle with the trunk. Thus conifers have a difficult time growing in glaze ice zones.
EVERGREEN COMPROMISES Evergreens which grow inside the glaze ice area show rather drastic adaptations to the destructive effect of ice. These adaptations usually compromise the plant's struggle for sunlight. They have, as a rule one or more of the following characteristics:
1. Small size - as American holly {Ilex opaca}
2. tough hard wood - as the yellow pine {Pinus ecinata}. In areas out of glaze or high winds, there is usually a benefit from light stiff wood, which permits faster growth, lower weight aloft, and broader bases in soft ground to prevent toppling.
3. Short branches, especially at high levels where wind velocity is higher - as the fir {Abies balsamea}.
4. Very flexible wood as the eastern red cedar {Juniperous virginiana}.
5. Down sloping branches as the spruce {Picea puntgens}.
6. Branches do not make a narrow upward angle with the trunk [Metcalfe] as almost all glaze ice conifers.
7. Less foliage as loblolly pine (McKeller).
8. Have asymmetrical branches as the pitch pine {Pinus rigida} and most dicotyledon angiosperms.
The reasons for these adaptations are obvious except for the last. The asymmetrical limbs are related to the additional destructive effects of high winds during or after an ice storm. I suggest that the cold makes the wood more brittle. The ice tends to increase the stiffness, inertia, and brittleness of the branch. The ice can also create a notch effect, causing a great concentration of strain where the ice cracks. Therefore, accentuated by the above circumstances, when the limb or tree is of such a length that it is in rhythm with the period of wind gust, the forces can be extreme [Geiger, p302]. If the ice is followed by high wind, the whole tree is liable to break off at the trunk [Bennett] or be uprooted. One good way to brake the force of rhythmical oscillations is to make use of asymmetrical limbs. If each branch is a different length and shape, and projects at a different angle, they tend to cancel out each other's oscillations because of different periods and directions of motions. Thus the large branches and trunk will be protected from extreme forces. Such an adaptation would be reinforced by hurricane winds and is characteristic of most southeast USA trees. Angiosperms are completely defoliated by hurricanes (Loope) which is an obvious advantage. One can easily see this phenomenon in action by observing any long limbed glaze ice tree during a wind and taking note that the limbs move in different directions at the same time. It is especially easy to see on leafless trees. This adaptation should not compromise competition for light much, and is universally adopted by glaze ice trees. It is especially strongly developed in angiosperms.
Short branches are obviously less efficient at competing for light, especially in lower latitudes. Nevertheless such branches are necessary on fully leafed trees because the ice load is approximately proportional to the square of the length of the limb while the leverage of the center of gravity is near to proportional to the length. In spite of this compromise, whole stands of white pines can be toppled by high winds {145 km per hour} and ice [Illick]. At high latitudes the advantage of long branches is much reduced because the light comes in at an angle so that where the foliage is sparse height of the tree starts to become a major consideration and limbs tend to be short.
Flexible wood would not seem to be a serious disadvantage in competing for light at first thought. However when an ice storm bends a red cedar or a birch tree to the ground, the flexible wood saves the tree's life, but these trees can not straighten back up again when the ice melts. Thus in the following spring they find themselves near the forest floor where light is blocked by stiffer trees. This is the most common kind of damage to trees which have under 5 cm. of trunk [Downs], and can affect many small trees in some storms.
Down sloping branches can form ice bridges which can reach the ground. Branches which grow at right angles to the trunk are less likely to split off than branches which grow up at narrow angles. In order to appreciate how seriously down sloping branches compromise the struggle for light as compared to branches which grow up at a sharp angle, it is instructive to analyze the light subtending power of a shape of a cone standing on its point {analogous to an elm}, as opposed to a cone standing on its base {analogous to a spruce}. This compares #5 to the negative of #6 in the previous list. It is the most serious compromise of all, so that it is pertinent to analyze the effect mathematically. Consider a forest in which for ease of analysis the following assumptions are made:
1. The trees compared are as high as the canopy which Will be taken as 100 feet {30.5 meters}. Feet are used because such a unit permits a believable canopy height to yield simple decimal numbers.
2. The light is absorbed at a uniform rate throughout, and the rate will be arbitrarily set at 5% per foot. In actual forests the rate is probably highest a couple of meters below the crown.
3. The plants use light at all intensities down to the forest floor. If light is not used near the floor, it will make the ratios obtained too small, not too large.
4. The average light intensity at the canopy top equals the symbol Io.
5. The trees compared form identical cones, compared as above.
6. The foliage is uniformly dense both vertically and horizontally.
These are assumptions which might reasonably be expected to approximate in some dense deciduous forests in summer. The mathematics are much simpler if the coordinates are made to start at the top of the canopy and are measured down. Call the distance from the top L.
Then according to Bouger's {Lambert's} law, Io over I=e to the KL
where I is the intensity of light at level L, and K is a constant, 0.05117 at 5% absorption.
From this we get: I=Io times e to the -KL
The volume of a cone standing on its base is given by: pie tan squared one half theta times the integration of (100 - L) squared dL, from 0 to L.
where theta is the angle of the cross section of the cone's peak and "integration of" is the symbol of integration.
The volume of a cone standing on its point is given by: pie tan squared one half theta times the integration of (100-L) squared dL, from 0 to L.
Now, if the horizontal increments of volume are multiplied by the light I and the fraction of light absorbed by a cubic foot of volume, the amount of light absorbed by a conical tree standing on its base is given by:
CIo pie tan squared one half theta times the integration of e to the -KL times L squared dL, from 0 to L, where C is the fraction of light absorbed per unit volume and is approximately equal to 0.5
If this integral is solved it becomes, ignoring the constants C, Io, pie, and tan squared one half theta:
e to the -KL times (L squared over K + 2L over K squared + 2 over K cubed) from L=0 to L=100.
Now if the above expressions are substituted for the full depth, we get 14.9 for the shape analogous to the spruce, and 291 for the shape analogous to the elm. These numbers are proportional to the light absorbed. The elm is thus 19.5 times more efficient at commandeering light than the spruce. With an advantage like that, the elm could easily afford to lose a branch or two every tenth winter or so if all it had to compete with were spruce. However, low absorption in spring or late fall would tend to decrease the above advantage in a real situation, the decrease being especially great where the growing season is short or winter photosynthesis is possible.
Of course dry or infertile conditions would also decrease the advantage if such conditions resulted in more sparse cover. Also it is implied that the light is coming from overhead. In Arctic regions much of the advantage would disappear. A much lower absorption value would be more typical in open pine barren forest, and here the ratio for the two cones is closer to one. Evergreen trees in the form primarily of pines encroach well inside the glaze ice. Even here trees with down sloping branches are not to be seen. Upward sloping branches only on deciduous trees are characteristic of the glaze ice zone. The upward angle averages around 45 degrees in Tennessee and about 30 degrees in Ohio. The most notable exception is the largely 80 degrees for only those branches which retain dead leaves on lower branches until early spring on some species common in Michigan. In Northern New Jersey right angle branches are fairly common. Perhaps this is because high rainfall makes possible the extra cellulose necessary for this less efficient way of gaining altitude. Some trees have evolved clever geometry of growth which ameliorates to a small extent the disadvantage of down sloping branches. Some conifers have branches which start out downwards and then curve up. Others have down sloping main branches on which the small branches grow up.
CONCLUSIONS
The angiosperms, from which most deciduous vegetation is derived, first appeared in the fossil record fully developed in the early Cretaceous at the 40th parallel near Korea [Basinger]. They were small leafed shrubs and herbs in seasonally arid habitats (Hickey), primarily along the banks of streams and in swamps [Retallack 1990] as ground cover under conifers. Many had a modern appearance [Axelrod 1952, p31], and were related to present day temperate forms such as sassafras, sycamore, magnolia, walnut, willow, poplar, tulip, birch, and persimmon by the time they reached North America, but had entered or been already in subtropical areas [Axelrod 1966]. About 35 million years later (110 million years ago they began to increase their share of the vegetation at a high rate in all climate zones from the equator to 60 degrees north as determined by fossil pollen, especially dicotyledon plants (Crane). The first angiosperms were strongest near the equator. Nevertheless I suspect that they initially had evolved some of their ability to use declining light to trigger physiological processes and perhaps even the excision layer in a higher latitude region in late Permian. It does not seem likely that a physiological process which requires declining light to trigger it, is likely to have initially started near the equator, so it must have at least started in the subtropics or higher. It would be virtually impossible that such extremely complicated features could have evolved from tropical progenitors [Axelrod 1952, p34] or any other progenitors in a short time later than from early Jurassic at least, and probably from Permian [Axelrod 1952, p32]. To have evolved so many families and genera with so many ubiquitous complicated processes, processes some of which arose in no other order of plants, would have required many millions of years.
This is because for long lived, large sized, and wind pollinated vegetation, evolution should be extremely slow.
For evolution to proceed rapidly a large population is necessary in order to increase the statistical chance of mutations and favorable recombinations [Mayr, p 585] and to decrease the chance of genes becoming extinct [Li]. The land can not support as large a population of large individuals as small. The length of time a species takes to reach reproductive age also affects speed of evolution in so far as chance favorable recombinations are concerned. A tree which takes 50 years to reach reproductive age, and then sends out seeds for another 100 years would have a chance of leaving a recombined descendant somewhere about once every hundred years on the average. It would be recombining one hundred times as slowly as an equal number of annuals for traits advantageous to mature plants, which would usually include speciation. It is conceivable to envision some trees on a stable area evolving one hundred thousand times as slowly as some annuals on an equal sized yearly disturbed area, since some annuals can easily occupy a thousand times less space. Since the first angiosperms were shrub sized or less [Hickey], the disparity relative to annuals or even perennial herbs was presumably less than the above, but they still must have required a considerable prior history, especially if the shrubs were only an advanced wave of migration from a forest containing plants which averaged larger. Permissible diversity is also an important factor in speciation. If wind pollinated trees are forced to make dense stands, speciation would be much slower than for insect pollinated species which can enter areas where its population density would be too low were it wind pollinated and thus have an increased chance of being cut off in a cul-de- sac as well as increasing the circumstances which would constitute a cul- de-sac. Insect pollination became common in mid Cretaceous [Bassinger], possibly because of increasing dominance of tropical rain forests by angiosperms. There is no possibility at all for a sudden evolution of progenitors of glaze ice angiosperm deciduous trees in early Cretaceous from an unknown ancestor, most of which trees fit into the above slowing categories, in only a few thousand years, or even a few million. It is possible that they evolved in an unknown tropical monsoon region [Axelrod (1966)] during Jurassic, but more probably as early as the Permian [Axelrod 1952, p57], possibly originally from rain forest trees [Whitehead]. However, the first angiosperms appearing on the fossil record were small leafed shrubs in seasonally arid habitats [Hickey].
Rate of gene change of chloroplast DNA seems to place the divergence of monocotyledon angiosperms (palms and grasses) from dicotyledon (broad leaved) at 200 million years or late Triassic [Wolfe & Gouy,et al 1989]. Phytochrome gene changes seems to place that divergence at 260 million years, or late Permian [Kolukisaoghi]. Chloroplast glyceraldehyde-3-phosphate dehydrogenase seems to place that divergence 320 million years ago, or all the way back to the Pennsylvanian [Martin et al]. Isocarbinol has been found in the Permian [Haike]. Since isocarbinol is found only in angiosperms, this would hint at precursors of angiosperms being present at least by the Permian. Any of these early estimates are infinitely more plausible than a Cretaceous sudden evolution. If so, progenitors of angiosperms go back at least to a time prior to the Triassic. Only groups which have no scaliform pitting at all, even in the metaxylem, can be eliminated as precursors. Coniferales, Ginkoales, and Gnetales are such eliminated plants (Hickey). Some think that seed ferns are the most probable source. Dicroidium "seed ferns" (glossopterids) were deciduous [Taylor][Gould] in the Permian and Triassic Antarctica [Taylor]. I consider it quite plausible that some of the most significant attributes, such as growth rings, could have evolved in the shadow of Carboniferous glaciers since woody plants with growth rings had become dominant by late Mississippian [Spicer] , and even a primitive excision leaf stem layer could have materialized then in a different group or in a cool Permian area even before their reproductive style differentiated. An indication that this last is at least plausible is that Glossopteric deciduous leaves are alleged to have been discovered on an ancient Gondwana forest floor in Antarctica (Pigg, p513), as well as a suspicion of leaf drop from Pteridosperms and deciduous Ginkgoales in early Cretaceous Antarctic [Douglas], and so progenitors of angiosperms could also conceivably have developed a deciduous habit this early, for if these orders could have an advantage from such a strategy another could as well. If it did happen this way it is not likely that Antarctica could have been the place where they subsequently did the bulk of their evolution because there were very few angiosperms in lower Cretaceous Antarctica [Douglas]. Deciduous genes could even have moved back and forth across all climate zones many times, since these attributes are useful for other purposes than cold adaptation.
I suspect that the initial angiosperm evolution was on the Ontong Java plateau, from which they migrated across Pacific Ocean atolls to the mainland in early Cretaceous.
You may see the concept above developed in an article in The General Science Journal shown in this URL; http://gsjournal.net/Science-Journals/Research%20Papers-Paleontology/Download/4719 .
QUOTES;
>It isn't what we don't know that gives us trouble,...it's what we know for sure,... that ain't so. Will Rogers
REFERENCES are below
ACKNOWLEDGMENTS
I would like to express my appreciation to Edward Zoll for checking the mathematics for me, to the U.S. Forest Service for permission to use their map, to American Telegraph and Telephone for permission to use their glaze ice map, and to the U.S. Dept. of Commerce for references and very useful maps, last not used in this article.
SOME LINKS to SIMILAR ARTICLES
SOME BIOLOGICAL HYPOTHESES
To see scientific names for common trees and shrubs see this site.
Continue to Permian marine phosphorus as caused by amphibians, especially dragonflies or to Cretaceous marine phosphorus as caused by sheet erosion from runway building termites, or to the effect of runway builders and incompetent ants on the phosphorus of Cretaceous soils and vertebrate bones and teeth especially dinosaurs.
Cretaceous and Eocene marine phosphates are explained as arising from sheet erosion of termite runways in the article about termites affect on soil.. For more details of the termite effect in different areas see the termites' affect on soil around the Paleocene.
LINKS TO MARS GEOLOGY
---- The Canyons of Mars as Erosion by Rivers of Silicone Dust
----For a hypothesis that explains the large volcanoes of Mars and the bulges associated with them as the disruption from the antipode (opposite side of a sphere) of a huge meteorite or comet impact, see this site.
GLOBAL WARMING
Climate warming as caused by denudation of soil.
SOME LINKS TO ASTROPHYSICAL PHENOMENA
The Cause of the Characteristics of Quasars
The Cause of the Cosmological Red Shift
For some dramatic views of a a virtual travel to Mars and then to outer space, a trip which would take thousands of years even inside our own galaxy, but compressed into 12 minutes, see this site.
SOME HEALTH ARTICLES
-- V. Electrolyte regulation (sodium and potassium) -- VI. Purpose of cortisol -- VII. Copper nutrition and physiology -- VIII. Availability of a book about potassium nutrition and physiology, especially as pertaining to heart disease, rheumatoid arthritis, gout, diabetes, high blood pressure, and hyperkalemia. --
Strategies for Chronic fatigue syndrome (CFS) and fibromyalgia -- When blood potassium is too high
Fluoride in city water will cause fluorosis discoloration of teeth, weakened bones, damage to the kidneys and immune system, bone cancer, and, worst of all, damage to the nerves resembling Alzheimers disease.
The health of people in the USA is abysmal and a major part of it is poor nutrition. As the 12th century physician, trying to cure by diet before he administers drugs, said; No illness that can be treated by diet should be treated by any other means" or as Hippocrates expressed it in 460 - 377BC; "If we could give every individual the right amount of nourishment and exercise, not too little and not too much, we would have found the safest way to health." It would seem that a healthy life style has been known for a long time. It is my belief that an unprocessed, unfrozen, not canned, high in vegetables diet would keep a large majority of people reasonably healthy and without the need for I would suggest that a partial solution to the problem of poor potassium nutrition would be to place a tax on all food that has had potassium removed by food processors and completely fund all Medicare and workmans compensation for injuries and disease that relate to rheumatoid arthritis, heart disease, and high blood pressure. This would also take the onerous tax burden now incurred for them and place it on the shoulders of those who cause the problem
There is an
If you have tried tumor necrosis factor or interferon to cure a cancer without success, perhaps an opioid antagonist drug called Naltrexone (Naltrexone in the large 50mg size, originally manufactured by DuPont under the brand name ReVia, is now sold by Mallinckrodt as Depade and by Barr Laboratories under the generic name naltrexone) that blocks some endorphin receptors. may work. Said blockage is thought to cause the body to temporarily secrete more endorphins, especially after midnight at night. These endorphins are thought to stimulate the immune system, and in particular to stimulate the TH-1 or type 1 antiviral response by decreased interleukin-4 and with increased gamma interferon and interleukin-2 and a simultaneous decrease of type 2 anti bacterial response [Sacerdote].
It appears to be especially effective for minimizing symptoms and retarding progression of multiple sclerosis (MS) (also see these sites hereand here and this site).
Low doses of Naltrexone (LDN), 1.5 to 4.5 milligrams, at bedtime is used (timing is important, and it is important not to buy slow release forms). It is said to have no known bad side effects at those doses other than stimulate the TH-1 or type 1 antiviral response by decreased interleukin-4 and with increased gamma interferon and interleukin-2 and a simultaneous decrease of type 2 anti bacterial response [Sacerdote]. There are also reports from an extensive survey in this site.
I think some clinical studies on Naltrexone are in order, and it should not be a prescription drug. Though side effects appear unlikely, it is not proven over longer periods. If you try it (it is a prescription medicine in the USA), it seems likely that you should discontinue if you get a bacterial infection in view of its inhibition of antibacterial response.
Olive leaf extract has shown clinical evidence of effectiveness against a wide range of viruses, including AIDS [Bihari], herpes, and cold viruses. It sometimes produces a Herxheimer or pathogen die off symptoms (from effectiveness against bacteria?). There is evidence that it is synergistic (reinforce each other) with Naltrexone. There have been a few case histories of improvement in what were probably arthritis patients and CFIDS patients. The active ingredient is said to be oleuropein or enolate. There has been very little follow up research done on it.
. Also it has been found that curcumin in turmeric or curry powder will inhibit several forms of cancer, including melanoma. People who live in India where these spices are eaten, have one tenth the cancer elsewhere.
Here is an article with anecdotal evidence for pressurized oxygen, zinc, vitamin B6, and vitamin C after head injuries. They also claim a fair percentage of prison inmates from psychiatric disorders after head injuries.
Taurine, or 2-aminoethanesulfonic acid, is an acidic chemical substance sulfonated rather than carboxylated found in high abundance in the tissues of many animals (metazoa), especially sea animals. Taurine is also found in plants, fungi, and some bacterial species, but in far less abundance. It is an amine with a sulfonic acid functional group, but it is not an amino acid in the biological sense, not being one of the twenty protein-forming compounds encoded by the universal genetic code. Small polypeptides have been identified as containing taurine, but to date there has been no report of a transfer RNA that is specifically charged with taurine [from Wikipedia]. It is essential to babies.
It has been found that supplements of the amino acid, taurine, will restore the abnormal electrocardiogram present during a potassium deficiency by an unknown mechanism. This information has been used in several case histories by George Eby to control a long standing type of cardiac arrhythmia called pre atrial contractions (PACs), a benign but irritating and nerve racking heart problem, with 2.5 grams of taurine with each meal.
Taurine is said to be low in the diets of vegetarians. The 2,500 milligrams recommended by the American Heart Association causes diarrhea in some people and should probably be reduced in those people. As much as 6 grams per day are said to be harmless [McCarty 2006, p67]. Taurine may make a copper deficiency worse, based on a single case history [Brien Quirk, private communication]. This may be because taurine may be mobilizing copper and zinc into the plasma [Li]. So if you should decide to take taurine, make sure your copper intake is at least adequate, as well as your zinc. Taurine has been used
for high blood pressure (it lowers borderline hypertension by decreasing epinephrine, but has no affect on normal tension) [Fujita], migraine headache (I suspect that less than 1000 milligrams can remove the headache caused by allergy to peanuts), high cholesterol, epilepsy, macular degeneration, Alzheimers disease, liver disorders, alcoholism, and cystic fibrosis, and depression. Keep in mind that some people may have a genetic defect that limits the amount of taurine tolerated and that adequate molybdenum may desirable. It is usually obtainable in health food stores.
A site is available which shows. foods which are high in one nutrient and low in another (including calories). This last site should be especially useful for a quick list of foods to consider first, or for those who must restrict another nutrient because of a genetic difficulty with absorption or utilization
If you use medication, you may see technical evaluations and cautions of drugs at the bottom of this site. The very extensive USDA Handbook #8 may be seen here. To access the information you must press "enter" to search, and then divide Kcal into milligrams of potassium. This last table is very comprehensive, is used in search mode, and even lists the amino acids. There are also links in it to PDF types of printouts from the table for individual nutrients available here Just click on the A or W button for the nutrient you desire. A table that has already done the potassium calculation is here in descending concentration or in alphabetical order.
There is a free browser called
Firefox, which is said to be less susceptible to viruses or crashes, has many interesting features, imports information from Iexplore while leaving Iexplore intact. You can also install their emailer. A feature that lists all the URLs on a viewed site can be useful when working on your own site. REFERENCES
Abell CA 1934 Influence of glaze ice storms upon hardwood forests in the
southern Appalachians. J. For. 32: 35-37.
Abbink OA 1998 Palynological investigations in the Jurassic of the North
Sea region. LPP contributions series No. 8, LPP Foundation, Utrecht.
pages 60, 159 plate XIII, figs d-g.(reference examined by N de Jersey).
Aerts R 1995 The advantages of being evergreen. Ecology and Evolution
10: 402-406.
Axelrod DI 1952 A theory of angiosperm evolution. Evolution 6: 29..
Axelrod DI 1966 Origin of deciduous and evergreen habits in temperate
forests. Evolution 20: 1.
Bassinger JF, Dilcher DL 1984 Poleward migration of early angiosperm
flora. Science 224: 511-513.
Bennett I 1959 Glaze, its meteorology and climatology, geographical
distribution. Science 224: 511.
Braun EL 1972 Deciduous forests of eastern North America. Hafner Pub.,NY.
Carlquist 1965 Island Life Natural History Press, Garden City Press.
Cornet, B 1989 Late Triassic angiosperm-like pollen from the Richmond
Rift Basin of Virginia, USA. Palaeontographica Beitrage zur
Naturgeschichte der Vorzeit 213 B: 37-87.
Cornet, B 1993 Dicot-like leaf and flowers from he late Triassic
tropical Newark supergroup riftzone, USA. Modern Geology 19; 81-99.
Crane PR Lidgard S 1989 Angiosperm diversification and paleolatitudinal
gradients in Cretaceous. Science 246: 675-8.
Darlington PJ,Jr. 1965 Biogeography of the Southern end of the World.
Harvard Univ. Press, Cambridge, Mass.
Dejersey NJ Grantmackie JA 1989 Palynofloras from the Permian, Triassic,
and Jurassic of New Caledonia. New Zealand J Geology & Geophysics 32:
463-476.
Doi H Takahashi M 2008 Latitudinal patterns in the phonological responses of leaf colouring and leaf fall to climate change in Japan.
Douglas JG & Williams GE 1982 Southern polar forests: the early
Cretaceous floras of Victoria and their paleoclimatic significance.
Paleogeogr., Paleoclimat., Paleoecology 39: 171-185.
Downs AA 1938 Glaze damage in the birch-beech-maple-hemlock type of
Pennsylvania & NY forest types. J. For. 36: 63-70.
Fosberg MA. Private communication.
Fretwell SD. Private communication.
Furomoto AS Webb JP Odegard ME Hussong DM 1976 Seismic 34: 71-90.
Geiger R 1950 The Climate Near the Ground, Stewart NM, translator.
Harvard University Press.
Haike V, Adam P, Trendel JM, Albrecht P, Schwark L, Vliex M, Hagemann H,
& Puttmann W.Isocarbanol through geological times: evidence for its
presence in the Permian and Triassic. Organic Geochemistry 23: -93.
Hill GF 1921 The white ant pest in northern Australia. Bull. No. 21
Commonwealth of Australia; Melbourne, Australia.
Hogan ME Shulz MW Slaytor M Cozolii RT Obrien RW 1988 Components of
termite and protozoal cellulases from the lower termite, Coptotermes
lacteus Froggatt. Insect Biochem 18: 45-51.
Hussong DM Wippeman LK Kroenke LW 1979 The crustal structure of the
Ontong Java and Manihiki Oceanic Plateaus. J. Geophys. Res. 84,
6003-6010.
Illick JS 1916 A destructive snow and ice storm. Forest Leaves 15: 10.
Kalshoven, LGE Observations on the nests of initial colonies of
Neotermes tectonae Damm in teak trees. Insects Sociaux 6: 231-242.
Kelly SRA Moncrieff ACM 1992 Marine molluscan constraints on the age of
Cretaceous fossil forests of Alexander Island, Antarctica. Geol. Mag.
129: 771-778.
Kolukisaoghi HU, Marx S, Wiegman C, Hanelt S, Schneider-Poetsch HAW 1995
Divergence of the phytochrome gene family predates angiosperm evolution
etc Selaginella and Equisetum arose prior to Psilotum. J. Molecular
Evolution 41: 329-337.
Kroenke LW Resig JM Leskie RM 1990 Hiatus and tephro chronology of the
Ontong Java Plateau: etc. Proceedings of the ocean drilling program
V130. NSF Joint Oceanographic Institutions.
Larson RL 1991 Latest pulse of earth:evidence for a mid-Cretaceous
superplume. Geology 19: 47-550.
Lawrence DB 1939 Some features of the Columbia River gorge with special
reference to asymmetry in forest trees. Ecol. Monog. 9:217.
Lee KE Wood TG 1971 Termites and Soils. Academic Press, London & NY.
Lemon PC 1961 Forest ecology of ice storms. Bull. Torrey Bot. Club 88:
21.
Li CC 1976 First Course in Population Genetics, p553. Boxwood Press,
Pacific Grove CA.
Loope L Duever M Herndon A Snyder J Jansen J 1994 Hurricane impact on
uplands and freshwater swamp forest. Bio. Science 44: 238-246.
Maycock PF 1962 Phytosociology of the deciduous forests of extreme
southern Ontario. Canadian J. Botany 41: 379.
Martin W, Gierl A, Saedler H 1989 Molecular evidence for pre-Cretaceous
angiosperm origins. Nature 339; 46-48.
Mayr E 1963 Animal Species and Evolution. The Belknap Press of Harvard
University Press, Cambridge.
McKeller AD 1942 Ice damage to slash pine, longleaf pine, and loblolly
pine plantations in the Piedmont section of Georgia. J. For. 40: 794-98.
Melville R 1966 Continental drift, Mesozoic continents and the
migrations of the angiosperms. Nature 211: 116.
Metcalfe EP 1959 Ice damage to trees. Weatherwise Mag. {whole issue
devoted to glaze ice}.
Michelle WA Phillips TL Peppers RA. 1985 The influence of climate and
depositional environment on the distribution and evolution of
Pennsylvanian coal swamp plants in: Tiffney BG, ed. Geologic Factors and
the Evolution of Plants. Yale Univ. Press,New Haven.
CD Monk 1966 An ecological study of hardwood swamps in south central Florida. Ecology 47; 649-654.
Monk CD 1966 An ecological significance of evergreeness. Ecology 47:
504-505.
Niering WA 1953 The past and present vegetation of High Point State Park
NJ. Ecological Monographs 23: 127-148.
Nimlos TJ, private communication.
Nur A Ben-Avraham Z 1982 Oceanic plateaus, the fragmentation of
continents, and mountain building. J. Geophys. Res. 87: 3644-3661.
Paramonov SJ 1960 Lord Howe Island, a riddle of the Pacific. Part II.
Pacific Science 14: 75-85.
Paramonov SJ 1963 Lord Howe Island, a riddle of the Pacific, Part III.
Pacific Science 17: 361-373.
Parish JT Spicer RA 1988 Late Cretaceous terrestrial vegetation: a near polar temperature curve. Geology 16;22.
Pigg KB Thomas NT 1993 Anatomically preserved Glossopteris stems with
attached leaves from the Central Antarctic Mountains, Antarctica. Am. J.
Botany 80: 500-516.
Pruitt WO Jr. 1960 Animals in the snow. Sci. Am. 202: 61.
Richards JH 1984 Ecophysiological characteristics of the winter
environment in establishment and tending of subalpine forest: research
and management. Proc. 2nd International Workshop Iufro Project Group Pl.
07-00, 3-5 Sept. 1984 Reideralp Switz: Turner H Tranquillini (eds).
{from abstract}
Retallack GJ 1990 Soils of the Past. Unwin Hyman, London.
Retallack GJ 1977 Reconstructing Triassic vegetation of eastern
Australia: a new approach for the biostratigraphy of Gondwanaland.
Alcheringa 1: 247-277.
Retallack G Dilcher DL 1981 Arguments for a glossopterid ancestry of
angiosperms. 7: 54-67.
Rodriguez, M., de Langre, E., Moulia, B., 2008. A scaling law for the effects of architecture and allometry on tree vibration modes suggests a biological tuning to modal compartmentalization. American Journal of Botany 95, 1523-1537.
Rohl U 1996 Aptian-Albian sea level history from guyots in the Western
Pacific. Paleoceanography 11: 595-624.
Sellier, D., Fourcaud, T., 2009. Crown structure and wood properties : Influence on tree sway and response to high winds. American Journal of Botany 96, 885-896
Spatz, H.C., Bruechert, F., 2000. Basic biomechanics of self-supporting plants: wind loads and gravitational loads on a Norway spruce tree. Forest Ecology and Management 135, 33-44.
.Spicer RA Parish JT 1986 Paeobotanical evidence for cool north polar
climates in middle Cretaceous (Albian-Cenomanian) time. Geology 14:
703-706.
Spicer RA Chapman JL 1990 Climate change and the evolution of
high-latitude terrestrial vegetation and floras. trends Evolution 5:
279-284.
Stoll, HM Schrag DP 1996 Evidence for glacial control of rapid sea level
changes in the early Cretaceous. Science 272: 1771-1774.
Sun, G., Dilcher, D.L., Zheng, Z., and Zhou, Z., 1998. In Search of the First Flower: A Jurassic Angiosperm, Archaefructus, from Northeast China. Science 282: 1692-1695.
Sun, G., Qiang, J., Dilcher, D.L., Zheng, S., Nixon, K.C., and Wang, X., 2002. Archaefructaceae, a new basal angiosperm family. Science 296: 899-904.
Taylor EL 1996 Enigmatic gymnosperms-structurally preserved Permian and
Triassic seed fens from Antarctica. Review of Paleobotany and Palynology
90: 303-318.
Tarduno JA, Sliter WV, Kroenke L, Leckie M, Meyer H, Mahoney JJ,
Telewski, F.W., Jaffe, M.J., 1986. Thigmomorphogenesis: Field and laboratory studies of Abies fraseri in response to wind or mechanical perturbation. Physiologia Plantarum 66, 211-218.
Musgrave R, Storey M, Winterer EL. 1991 Science 254: 399.
Wade, J.E., Hewson, E.W., 1979. Trees as a local climatic wind indicator. Journal of Applied Meteorology 18, 1182-1187.
Waring RH FRanklin JF 1979 Evergreen coniferous forest of the Pacific
Northwest. Science 204: 1380.
Weber CE 1993 Cretaceous termites and soil phosphorus. Indian J. Soil
Biol. Ecol. 13: 108-121.
Whitehead DR 1969 Wind pollination in the angiosperms: Evolutionary and
environmental considerations. Evolution; 23: 28-35.
Wolfe, KH, Gouy, M, Yang, YW, Sharp, PM, & Li,WH 1989 Date of
monocot-dicot divergence estimated from chloroplast DNA sequence data.
Proceedings National Academy Sciences 86: 6201-205.
Wolfe, JA & Upchurch GR 1986 Vegetation, climatic and floral changes at
the Cretaceous-Tertiary boundary. Nature 324: 148-151.
Wolfe JA 1987 Late Cretaceous-Cenozoic history of deciduousness and the
terminal Cretaceous event. Paleobiology 13: 215-226.
Wolfe JA 1987 An overview of the origins of the modern vegetation and the
flora of the northern Rocky Mountains. Annals of the Misouri Botanical
Gardens 74; 785-803.
. This article has been updated in March 2014
.
There is also an article which proposes some speculation about diabetes. There is a site that contains reviews of natural remedies for many diseases .
See this site for evidence of a correlation between magnesium deficiency and cancer.
This page has been visited times